L'œil est un système formateur d'image. Il est souvent comparé à un objectif photographique qui, correctement mis au point, donne d'un objet une image renversée située dans le plan du film. L'analogie est discutable, car les différences sont multiples. Toutefois les plus importantes ne résident pas dans les systèmes optiques proprement dits : milieux transparents, mise au point, image plane ou courbe..., mais dans les détecteurs et la façon dont ils transforment l'image qui se forme sur eux. Qu'ils soient détecteurs argentiques (émulsion classique), ou électroniques (capteurs CCD des appareils numériques), le traitement de l'image optique qu'ils opèrent est sans commune mesure avec la complexité du traitement de l'image rétinienne qu'effectuent la rétine, les voies optiques et le cortex visuel. Comprendre la couleur, et dépasser les conflits agitant physiciens, physiologistes, psychologues et artistes, qui utilisent le même mot, dans des champs d'investigation différents, nécessite de prêter quelque peu attention au mécanisme de l'élaboration de ce signal coloré, jusqu'à sa confrontation, en fin de parcours, aux représentations et interprétations mentales. C'est donc autour de la rétine, et de la neurophysiologie de la vision, qu'il faut rechercher, dans un premier temps, ce qui aurait pu - s'il était né un siècle plus tard, réconcilier peut être Goethe avec Newton. 1. La rétine en tranchesNotre rétine a la particularité de tourner le dos à la lumière et les cellules visuelles ne reçoivent qu'une lumière indirecte, retrodiffusée par l'épithélium pigmentaire.Si l'on assimile l'œil à une sphère dont le rayon interne serait d'environ 11,5 mm, la rétine tapisse à peu près les trois quart de cette sphère (1250 mm2), sur une épaisseur qui peut varier de 0,5 mm à 0,1 mm (centre fovea). Les détecteurs visuels associés à la vision des couleurs, à savoir les cônes, sont principalement regroupés sur la macula, de forme approximativement circulaire, et plus précisément sur la fovea, qui forme une dépression sur la rétine car le cablage neuronal est rabattu vers l'extérieur. Quelques ordres de grandeur :
* l'extériorisation représente l'ouverture angulaire du cône objet associé à la surface rétinienne considérée. 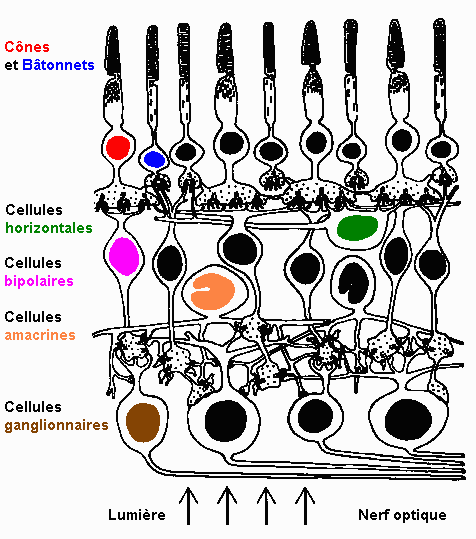 La structure de la rétine est représentée sur le schéma ci-dessus. Il apparaît deux structures de cellules organisées radialement (cônes et bâtonnets, bipolaires, ganglionnaires) ou transversalement (horizontales, amacrines). Dans la rétine la vision des couleurs se structure principalement en deux niveaux :
2. Le schéma de Walraven |
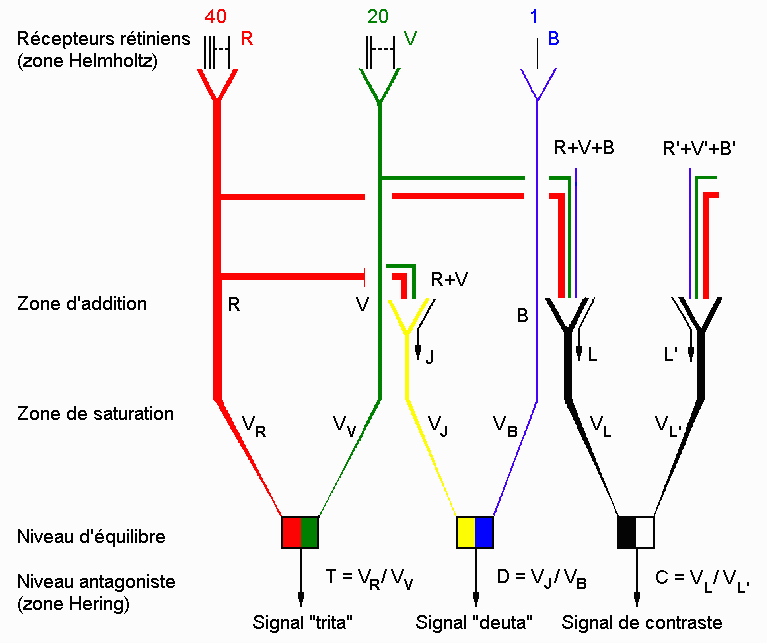
|
Les cônes S, M et L ne sont pas en quantité égale. Ainsi, pour 1 cône S (sensible au bleu), on trouve 20 cônes M (sensibles au vert) et 40 cônes L (sensibles au rouge). Dans le schéma qu'il a proposé en 1973, Walraven tient compte de cette distribution. La transformation du signal chromatique se fait ŕ différents niveaux, le premier étant celui de Helmholtz, et le dernier celui de Hering.
Mais... 3. Rien n'est jamais acquis...Les résultats énoncés plus haut ne sont qu'une étape, déjà dépassée, des recherches sur les mécanismes de la vision des couleurs. Car l'histoire de cette couleur et de la façon dont elle est modelée, se continue jusqu'aux aires corticales qui logent à l'arrière de notre crâne, en empruntant le chemin du nerf optique, du chiasma, des corps genouillés latéraux et des radiations optiques.Situer la réconciliation de Goethe et de Newton au niveau des cellules ganglionaires est donc quelque peu exagéré. Disons que c'est plutôt le lieu d'un armistice, et que le débat continue, sur d'autres fronts... On pourra pour approfondir cette complexité lire l'article d'Yves Galifret dans l'Encyclopædia Universalis [7], ou bien le texte de Peter Gouras, directement accessible sur le Net [10]. |
|||